And while there were also minimal impacts on loops, our PCMC data suggest that BAF and INO80C are not general regulators of chromatin looping but may instead fine-tune regulatory interactions at distinct key developmental loci in ES cells.
18.09.2025 15:00 — 👍 1 🔁 0 💬 0 📌 0
Overall, our study suggests that esBAF and INO80 have limited influence on large-scale 3D genome architecture. However, subcompartment organization is sensitive to remodeler depletion, indicating a more nuanced role for these complexes in shaping chromatin topology at sub-megabase scales.
18.09.2025 15:00 — 👍 1 🔁 0 💬 1 📌 0

Enhancer-promoter loops impacted by BRG1 or INO80 loss are enriched for OCT4, SOX2, NANOG and SUZ12, and are described as bivalent locations.
For both these remodelers, these tend to be OSN bound and also Suz12 bound, and enriched as bivalent promoters. Together, these data suggest that the enhancer-promoter loops esBAF promotes and INO80C restricts are pluripotency and bivalency related.
18.09.2025 15:00 — 👍 0 🔁 0 💬 1 📌 0

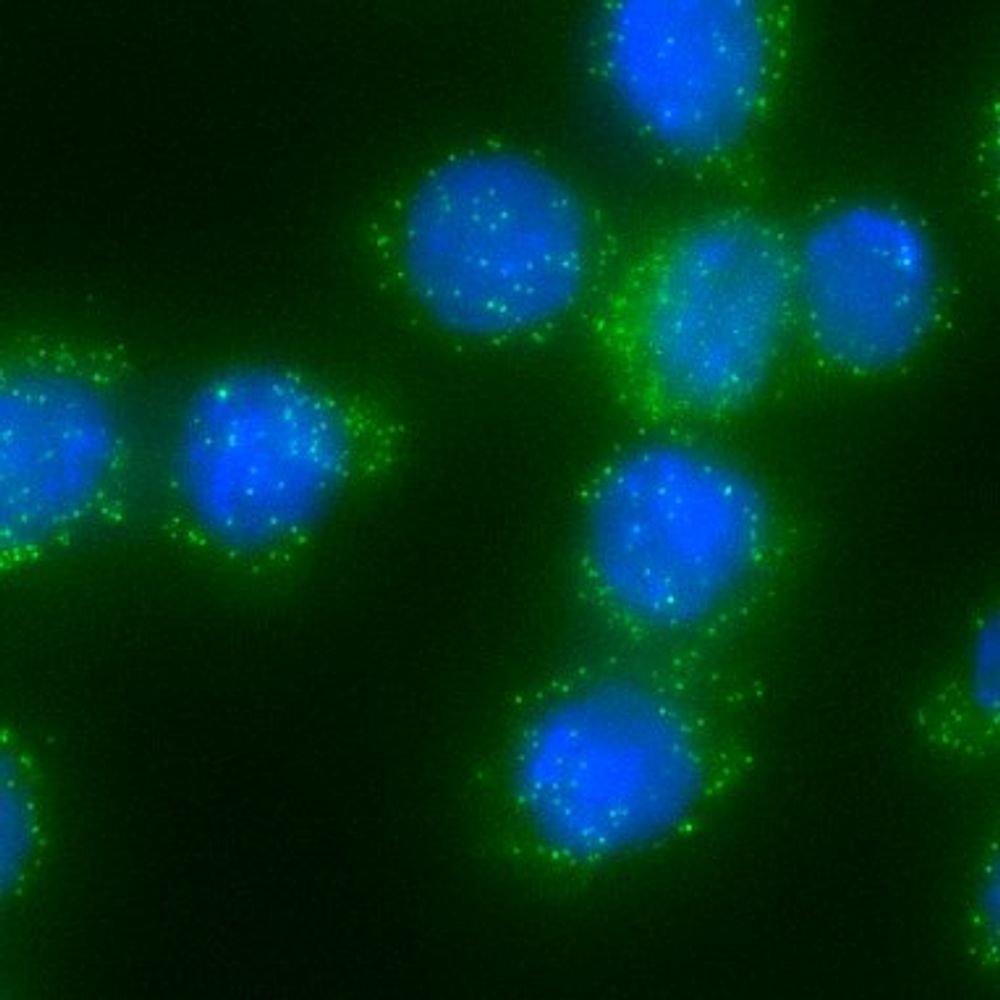
Promoter capture microC (PCMC) enriches for TSS-based loops and shows decreased loops in Brg1 KD and increased loops in Ino80 KD
Therefore, to investigate the impact of these remodelers more directly on enhancer-promoter loops, we performed promoter capture microC (PCMC) and found that KD of BAF ATPase BRG1 results in decreases in some E-P loops and KD of INO80C ATPase Ino80 results in increases in some E-P loops.
18.09.2025 15:00 — 👍 0 🔁 0 💬 1 📌 0

Our Hi-C data detects limited TSS-based loops, but is enriched for CTCF-based (architectural) loops
Although we sequenced our Hi-C datasets deeply, we had a tough time calling non-architectural loops (most loops were CTCF-based, and not as many were TSS-based)
18.09.2025 15:00 — 👍 1 🔁 0 💬 1 📌 0

Hi-C data analyzed for TAD structures
Similar to work from the Schubeler lab for BAF, we find that loss of neither BAF nor INO80C impact TAD structures significantly.
18.09.2025 15:00 — 👍 0 🔁 0 💬 1 📌 0
We therefore thing that BRG1 and INO80 contribute to the maintenance of active subcompartment organization (we are saying "fine-tune"), with effects that are especially pronounced at loci where these remodelers bind and/or their loss impacts transcription.
18.09.2025 15:00 — 👍 0 🔁 0 💬 1 📌 0

subcompartment analysis of Hi-C data showing impact of remodeler depletion on subcompartments
However, there is a reproducible effect on subcompartment structures, where upon KD of either ATPase, we observed a modest increase in the total amount of the genome assigned to inactive subcompartments, especially at locations that these remodelers bind and regulate transcription.
18.09.2025 15:00 — 👍 0 🔁 0 💬 1 📌 0

heatmaps from Hi-C data showing compartment structures
48hr KD of the ATPase subunit for either complex has almost not impact on compartments, assessed by Hi-C:
18.09.2025 15:00 — 👍 0 🔁 0 💬 1 📌 0
Oh please add me! Thank you for making this!
29.08.2025 11:21 — 👍 1 🔁 0 💬 1 📌 0
🧬 Transcription elongation by RNA polymerase II relies on a web of elongation factors. Our new work shows how IWS1 acts as a modular scaffold to stabilize & stimulate elongation. Fantastic work by Della Syau! www.biorxiv.org/content/10.1...
29.08.2025 10:14 — 👍 56 🔁 23 💬 1 📌 1
These studies describe a mechanism by which the Chd1 is coupled to transcription elongation and the molecular consequences when this coupling is disrupted and provide insight into a domain in Chd1 for which little is known.
28.08.2025 23:07 — 👍 0 🔁 0 💬 0 📌 0

the Rtf1 mutation not being sufficient to break the interaction, and/or represent the complexity and redundancy present in this more complicated system.
28.08.2025 23:07 — 👍 1 🔁 0 💬 1 📌 0

So in a murine cell line system (ES cells) what happens? We made the CHCT deletions and an Rtf1 mutation ES cell line, but did not observe the same nucleosome shifts. This may be due to reduced protein levels observed in the Chd1 and Chd2 CHCT deletion cell lines...
28.08.2025 23:07 — 👍 0 🔁 0 💬 1 📌 0

Chd1 and Rtf1 are conserved proteins, so we wanted to know if this interaction is conserved in mammalian systems. Y2H using murine constructs demonstrate an interaction between Chd1 and Rtf1 as well as the related remodeler, Chd2, and Rtf1. CHCT deletions and Rtf1 mutations reduce this interaction
28.08.2025 23:07 — 👍 0 🔁 0 💬 1 📌 0

Histone PTMs, K4me3 and K36me3 are also shifted 5'
28.08.2025 23:07 — 👍 0 🔁 0 💬 1 📌 0
Notably, these precise interaction mutants seem to have a greater defect in nucleosome localization relative to full deletion, which demonstrates the multifunctional aspect of these proteins as well as the need for precision mutations.
28.08.2025 23:07 — 👍 0 🔁 0 💬 1 📌 0

As Chd1 is an important nucleosome remodeler, what does this loss of appropriate localization mean to genic nucleosomes? Well, nucleosomes (and overlapping dinucleosomes aka hexasome-nucleosomes) are also shifted 5', and this is exacerbated in Isw1 delete.
28.08.2025 23:07 — 👍 0 🔁 0 💬 1 📌 0

By mutating either side of this interaction (deleting the CHCT domain or precise point mutations in Rtf1), we find that while Rtf1 occupancy is unaltered, Chd1 occupancy is shifted 5'
28.08.2025 23:07 — 👍 0 🔁 0 💬 1 📌 0

Building off the finding from the Arndt lab in 2003 (10.1093/emboj/cdg179), we narrowed down an interaction between the understudied Chd1 CHCT domain and a N terminal LALA box in Rtf1, a member of the Paf1 elongation complex, which form a direct interaction in budding yeast
28.08.2025 23:07 — 👍 0 🔁 0 💬 1 📌 0

if i were a better person i would do a tweetorial on our recent paper - but, hey, read the paper. its right there, waiting for you. #Giantvirus #histones are cool! their #nucleosomes are cool. check it out.! thats the tweetorial.
rdcu.be/exXBX
26.07.2025 19:08 — 👍 84 🔁 19 💬 1 📌 1
Oh gosh Anders I am so sorry
21.08.2025 11:25 — 👍 1 🔁 0 💬 0 📌 0

Tenure track position alert! Assistant Professor of Molecular Therapeutics in MCB @UCBerkeley. Apply and come be our colleague! 🎉
aprecruit.berkeley.edu/JPF05098
21.08.2025 04:00 — 👍 14 🔁 9 💬 1 📌 0
We are hiring! The Dept of Microbiology and Molecular Genetics is looking for faculty at the Assistant or Associate professor level (tenure track). Please consider joining our vibrant microbiology and immunology community at the University of Pittsburgh School of medicine
20.08.2025 19:41 — 👍 106 🔁 107 💬 1 📌 4
Congrats Marco!
21.08.2025 11:22 — 👍 1 🔁 0 💬 1 📌 0
Scientist & serial-punner I Chromatin dynamics & gene regulation in early development I MSc. @reNEW | Ph.D. @embl | 🇩🇰 🇮🇹 |
Biologist by training, basketball player at heart
Professor for Translational Epigenetics & Genome Architecture, University Medical Center Goettingen, Germany.
www.papantonislab.eu
assistant prof at uOttawa studying epigenetics and brain development
Group leader in the Genome Biology Dpt of I2BC. My team studies how loss of function mutations in chromatin remodelers lead to cancer.
Working @adelmanlab.bsky.social @hms-bcmp.bsky.social + Vik Khurana lab @mgbresearch.bsky.social. Previously at Eilers/Wolf Lab @uni-wuerzburg.de. Interested in transcription and dsRNA biology - with a special focus on RNA Pol II.
Genetics professor at Harvard Medical School. Interested in RNA life cycles and genome organization across the cell, from the nucleus to mitochondria.
Teif lab at the University of Essex. We work on gene regulation in chromatin and applications to liquid biopsies, using approaches of genomics, biophysics, bioinformatics & AI. Our focus is nucleosomics, TF binding, CTCF, cfDNA. https://generegulation.org
Group leader and Wellcome Sir Henry Dale Fellow based at the Centre for Human Genetics at Oxford University. Interested in genomics, single-cell technologies, 3D DNA folding and chromatin disruption in human disease. He/him.
Enhancers and 3D genome structure | Lecturer @uniofexeter.bsky.social | sites.google.com/view/flynnlab/
Group leader, transcription dynamics in single cells, NKI
Developmental biologist working at CNRS/IGMM/University of Montpellier, interested by gene regulation in drosophila embryos. we use quantitative imaging to monitor mRNA lifecycle.
Head of Genome Biology Dept @EMBL,
Scientist, Principal investigator, Professor
Exploring genome regulation during development,
and everything to do with enhancers.
3D genome, chromatin topology,
cell_fate, embryonic Development,
Single Cell genomics
The Nogales Lab @ UC Berkeley. #CryoEM #Transcription #Microtubules #ChromatinModification Bluesky account managed by lab members.
We study transcriptional regulation and chromosome folding using an interdisciplinary approach combining wet- and dry-lab methods.
https://giorgettilab.org
@fmiscience.bsky.social
Group leader at the Danish Cancer Institute and Professor at University of Copenhagen #NNFCPR. Interested in epigenetics, genome maintenance, aging & cancer, creativity & innovation, society in general. Opinions are my own.
Computational & Genomics Lab studying 3D genome function and dynamics @ Netherlands Cancer Institute
Graduate student @mtleelab.bsky.social U Pitt - Evolution of post-transcriptional gene regulation in animal early embryonic development
I am a physician-scientist and sarcoma medical oncologist. My lab studies how alterations in the epigenome influence cancer biology. Our goal is to use that knowledge to find better ways to help patients. nacevlab.com
Small RNA biologist at UPenn & CHOP
www.coninelab.com
Interested in small RNAs, germlines, inheritance, sports, and Shiba Inu’s