Why do we care if any of it matters in the grand scheme of things if the Dow is over 50,000?
14.02.2026 05:15 — 👍 0 🔁 0 💬 0 📌 0
I remain convinced that drivers who don't use turn signals lack a theory of mind.
12.02.2026 15:33 — 👍 2 🔁 0 💬 0 📌 0

Insightful commentary on the new Ileret Homo habilis material by Ian Tattersall: anatomypubs.onlinelibrary.wiley.com/doi/10.1002/...
11.02.2026 12:01 — 👍 5 🔁 1 💬 0 📌 0
Many congrats to all our pals up there in Fort Collins! Nice work.
11.02.2026 23:45 — 👍 1 🔁 0 💬 0 📌 0
Fantastic! I'm glad that worked out. Happy to help spread the word locally if helpful.
11.02.2026 19:25 — 👍 0 🔁 0 💬 0 📌 0

A graphic reading "AABA Art, Culture and Science Engagement Exhibition" with stylized DNA, skulls, and primate faces. Art by Michelle Bezanson
The Art/Culture/SciComm Expo returns for the 2026 @bioanth.org meeting! #Science -related #SciArt photography, poetry, needlepoint, painting, sculpture, games, posters about podcasts or local engagement work…all these and more are welcome! Submission deadline is Mar 7.
forms.gle/kk7qCfQGXe6h...
30.01.2026 19:27 — 👍 5 🔁 4 💬 0 📌 2
Were you and Kate able to identify any institutions here in Denver interested in an outreach session?
11.02.2026 17:58 — 👍 0 🔁 0 💬 1 📌 0

Figure 1 from Blasi-Toccacceli et al. Views of OMO VE3-10063. Humerus OMOVE3-10063a, proximal portion: A,lateral; B,anterior; C,medial, D,posterior; E,proximal. Humerus OMOVE3-10063a, diaphysis: F,lateral; G,anterior; H,medial;I,posterior. Humerus OMOVE3-10063a, distalportion: J, Anterior; K, posterior; L,distal.Clavicle OMOVE3-10063c: M, Proximal; N, posterior; O, distal; P,a nterior. Scapula OMO-VE 3-10063b: Q, Anterior; R, posterior; S, lateral. Cervical neural arch OMOVE3-10063d: T, Superior; U, inferior. Cervical body OMOVE3-10063e: V,Anterior; W,posterior;X,inferior;Y,superior. RibOMOVE3-10063f: Z-superior ; a,inferior. See the detailed description of the individual in the Supplementary Material (Supplementary Text “Description of OMO VE 3-10063”) and Table S1. Abbreviations: Prox.: proximal; Dist.: distal; Med.: medial; Lat.: lateral; Post.: posterior; Ant.: anterior.
Preprint describing an interesting 1.84 million-year-old partial hominin skeleton (mostly shoulder) from the Shungura Formation in Ethiopia. Unfortunately, no craniodental remains to confirm taxon, but it shows some derived features linking it to genus Homo.
paleo.peercommunityin.org/articles/rec...
11.02.2026 17:46 — 👍 6 🔁 1 💬 0 📌 0
Me too. I really miss working there.
09.02.2026 13:17 — 👍 1 🔁 0 💬 0 📌 0
Great analytical work done on the specimen by our collaborator Matteo Romandini.
09.02.2026 04:41 — 👍 2 🔁 0 💬 0 📌 0
A Mesolithic superb owl (Bubo bubo) from a beautiful site our crew excavated in northwestern Italy back in 2018.
Citation if you're interested to know more:
09.02.2026 04:34 — 👍 8 🔁 1 💬 1 📌 0
Oh damn. Brilliant. Didn't even occur to me.
09.02.2026 04:27 — 👍 1 🔁 0 💬 1 📌 0

AI is not a peer, so it can’t do peer review
If we still believe that science is a vocation grounded in argument, curiosity and care, we can’t delegate judgement to machines, says Akhil Bhardwaj
'to treat peer review as a throughput problem is to misunderstand what is at stake. Review is not simply a production stage in the research pipeline; it is one of the few remaining spaces where the scientific community talks to itself.' 1/3
03.02.2026 08:17 — 👍 363 🔁 154 💬 6 📌 21

This is figure 2, which shows MLP-3000-1 partial left mandible.
A paper in Nature reports the discovery of fossilized remains of an extinct hominin called Paranthropus in north-eastern Ethiopia, dating to about 2.6 million years ago, which expands the number of hominin species known to have been in the region at that time. go.nature.com/4r37ceu 🏺 🧪
02.02.2026 02:48 — 👍 42 🔁 12 💬 0 📌 1
It's not my research, but @annemiekemilks.bsky.social and @katharv.bsky.social certainly did some nice work!
01.02.2026 01:23 — 👍 2 🔁 0 💬 0 📌 0
I should mention...many thanks to @heatherfsmith.bsky.social for being a fantastic editor and making publication at The Anatomical Record as painless as can be. The journal is a great venue for this work.
29.01.2026 00:41 — 👍 3 🔁 0 💬 0 📌 0
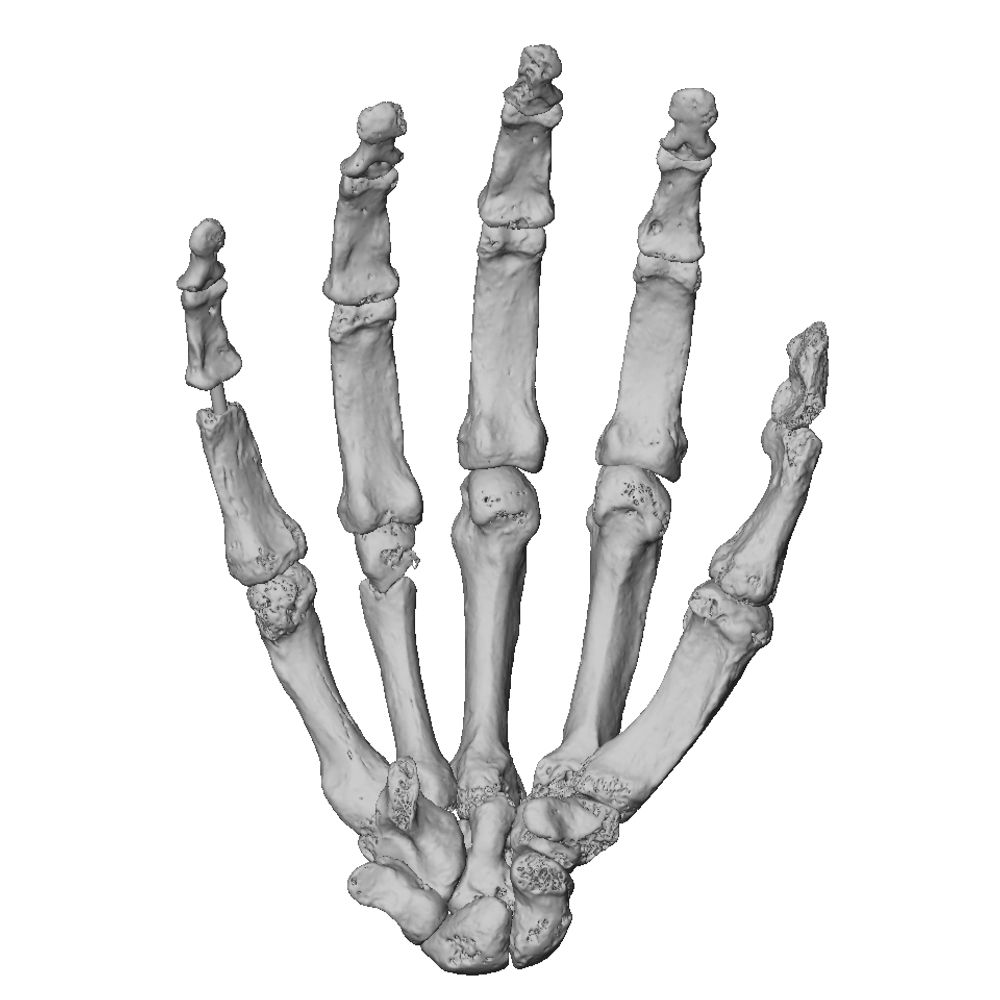
Publications / Orr et al Hominin postcranial remains from Drimolen Main Quarry South Africa | Human Fossil Record Archive
Publications / Orr et al Hominin postcranial remains from Drimolen Main Quarry South Africa
We've made our 3D scans of these Drimolen postcranial specimens available in the online archive at The Human Fossil Record. Hopefully access to these fossils contributes well to other studies of early hominin postcranial anatomy & variation. Meanwhile, fieldwork & analysis at Drimolen continue. 9/
29.01.2026 00:11 — 👍 13 🔁 4 💬 1 📌 0

Figure 2 from Orr et al 2026.
Arm and forearm elements including an adult distal humerus (a), proximal ulna probably from a subadult (b), fetal/perinatal humeri (c), and fetal/perinatal radii (d). Based on spatial context and developmental age, DNH 110 might be associated with DNH 112 (proximal tibia).
Another challenge w/ the Drimolen assemblage is a high frequency of young juveniles (several fetal/perinatal). Unfortunately, there aren't a lot of comparative data on fossils or even extant primates to facilitate anatomical analysis. But it's very interesting to have so many baby hominins. 8/
29.01.2026 00:11 — 👍 2 🔁 0 💬 1 📌 0
Despite taxonomic challenges, these Drimolen specimens combined w/ other recent insights into the early hominin postcranium suggest that the last common ancestor of Paranthropus & Homo may have had an increased commitment to terrestrial life, which has interesting paleobiological implications. 7/
29.01.2026 00:11 — 👍 2 🔁 0 💬 1 📌 0
Such comparative observations suggest that Paranthropus & Homo share derived locomotor anatomy, making taxonomy of the Drimolen specimens and others from South Africa (e.g., SKX 16699 from Swartkrans) rather ambiguous. 6/
29.01.2026 00:11 — 👍 2 🔁 0 💬 1 📌 0

Figure 15 from Orr et al 2026.
Comparative analyses of the DNH 117 nonhallucal pedal proximal phalanx. (a) Bivariate ln-ln plot of the midshaft area calculated as the mediolateral width (ML) × dorsoplantar height (DP) against interarticular length; sample includes recent H. sapiens (open triangles), Pan sp. (black plus signs), Gorilla sp. (open circles); Pleistocene H. sapiens (filled black triangles); Neandertals (open triangles); H. naledi (blue triangles); A. afarensis (red squares); P. boisei KNM-ER 101000 (green square); Swartkrans specimen SKX 16699 (green diamond), A. cf. africanus StW 355 (yellow square); DNH 117 (yellow star). Because ray is unknown for most of the fossil specimens, the extant sample combines all rays II–V. Least-squares regression line fit to full extant sample (ln midshaft area = 1.87 × ln midshaft area − 2.58). (b) Dorsal orientation of the metacarpophalangeal joint facet quantified as the articular angle following Trinkaus and Patel (2016) with comparative data from the same source; (c) longitudinal shaft curvature quantified as the included angle following Stern Jr. et al. (1995). (d) 3D surface renderings of DNH 117 and select comparative specimens in plantar view (top) and “lateral” view (bottom). Images of all specimens are scaled to the same proximodistal length. Plots b and c include the median of the sample along with the interquartile range (box) and the full range (whiskers).
An informative Drimolen piece is a toe bone (DNH 117) that has a robust, flat shaft with a dorsally oriented proximal joint suggesting humanlike toeoff during walking. Genus Homo shows this, but so does a foot of Paranthropus boisei from Kenya dated to 1.5 million years ago (KNM-ER 101000). 5/
29.01.2026 00:11 — 👍 3 🔁 0 💬 1 📌 0

Figure 10 from Orr et al 2026.
Shape analysis of the DNH 32 distal humerus. (a) reconstructed cross section of the distal shaft with landmarks (black dots) and semilandmarks (smaller red dots) on the elbow region. (b) Procrustes distances from the DNH 32 specimen to each of the fossil comparative specimens (with a lower value indicating a closer match)—filled symbols indicate fossils identified from a taxon-specific temporal/geographic context and numbers correspond to those in Supporting Information Table 1; and (c) plot of the first three principal components (PCs) for the geometric morphometric analysis of the elbow region. “Australopithecus (southern)” includes A. africanus and A. sediba specimens; “Australopithecus (eastern) includes A. anamensis and A. afarensis specimens. Paranthropus boisei specimens were included in the calculation of the principal component axes, but are not plotted for visual clarity of the figure. The convex hull associated with the purported P. boisei specimens falls with the other fossils along PC1 and overlaps that of Australopithecus (eastern) but no other fossil groups.
The taxonomy of most of these Drimolen specimens is unresolved, but we can likely assign a distal humerus (DNH 32) & partial pelvis (DNH 43) to Paranthropus, while two juvenile vertebrae are probably associated with a cranial vault of a young Homo aff. erectus individual (DNH 134). 4/
29.01.2026 00:11 — 👍 0 🔁 0 💬 1 📌 0
We don't have a lot of diagnostic features to identify Paranthropus vs. early Homo postcrania. Both were probably quite generalized & primitive in many ways (Australopithecus like) and some derived features are likely to be shared between the two genera because of their close relationship. 3/
29.01.2026 00:11 — 👍 3 🔁 0 💬 1 📌 0
Drimolen, like other sites in The Cradle of Humankind, has a mixed craniodental assemblage of Paranthropus robustus & early Homo. That makes sorting out the taxonomy of isolated postcrania difficult if not impossible. This is especially true because Paranthropus & Homo are probably sister taxa. 2/
29.01.2026 00:11 — 👍 2 🔁 0 💬 1 📌 0
Part of SUNY at Buffalo's Anthropology department, led by Dr. Noreen von Cramon-Taubadel. We study primate morphological variation and evolution!
---
https://bheml.weebly.com/
https://www.instagram.com/bhem_lab/
Archaeologist. PhD on MSA lithic technology. Looking for a postdoc 👀
Albacete -Burgos
News, events and research from the Department of Archaeology and the McDonald Institute for Archaeological Research, University of Cambridge
A/Prof Palaeolithic Archaeology, University of Oxford. Island archaeology; anthropology; human dispersal; zooarchaeology; lithics
NERC CREATES DLA PhD student, University of Cambridge | Human Evolution in Africa, Plio-Pleistocene, Palaeoanthropology, Archaeology | she/her
Director of Language & Genetics at Max Planck Institute, Nijmegen.
Tracing the complex connections between genes, brains, speech & language.
Website: https://www.mpi.nl/people/fisher-simon-e
ORCID: https://orcid.org/0000-0002-3132-1996
Visit https://human-fossil-record.org and browse our collections of digital files (screenshots, interactive surface models, and micro-CT stack movies) of fossil hominins and extant apes! Request of micro-CT data is also available for select collections.
Professor of Evolutionary Biology at Anglia Ruskin University, Cambridge.
www.thepeergroup.org.uk
PEER Group research is focused on primate evolution and ecology, with a particular emphasis on vocal communication and the evolution of language.
The podcast where scientists review movies about prehistoric people. https://screensofthestoneage.com
Enabling sustainable solutions for the long-term survival of the orangutan in the wild.
Find us at www.orangutanlandtrust.com!
Funded by the European Research Council (No. 949330). In collaboration with the FECYT– Ministerio de Ciencia, Innovación y Universidades
The Official BlueSky of the Department of Anthropology at Texas A&M University 👍 #tamuanth
Paleontologist, Research Fellow at @upvehu.bsky.social
The MATRIX project – Into the Sedimentary Matrix: Mapping the Replacement of Neanderthals by early Modern Humans using micro-contextualized biomolecules – is a European Research Council (ERC) starting grant attributed to Dr. Vera Aldeias.
Postdoc Researcher | Human evolution and anatomy|Long bones|Geometric Morphometrics | Biomechanics
Articles from Sounds Magazine 1975 - 1980
Archivist, Steve "Stig" Chivers.
Science journalist covering archaeology, paleontology, and Earth science | Words for Science, New Scientist, Science News, and Live Science 🧪🏺
Post doc @MPI_EVA Dept. of Human Origins
Leakey Foundation grantee 🐵 wild chimpanzee positional behaviour and the origins of bipedalism 🌳👣
Field blog 👉 https://apewanders.wordpress.com/
Stuttering Science Writer and occasionally charismatic megafauna.
Covering all things natural history at Smithsonian and elsewhere. Bylines NY Times, Scientific American, etc.
Amateur scientific illustrator
https://jacktamisiea.com/
 16.02.2026 02:32 — 👍 5 🔁 1 💬 0 📌 0
16.02.2026 02:32 — 👍 5 🔁 1 💬 0 📌 0