📢 Please help spread the word: We’re hiring a Head of Student & Postdoc Affairs to coordinate our international PhD program, provide guidance & career counseling, lead training programs, collaborate on EDI initiatives, and manage alumni relations. Apply at www.fmi.ch/education-ca...
04.12.2025 07:23 — 👍 2 🔁 6 💬 0 📌 1

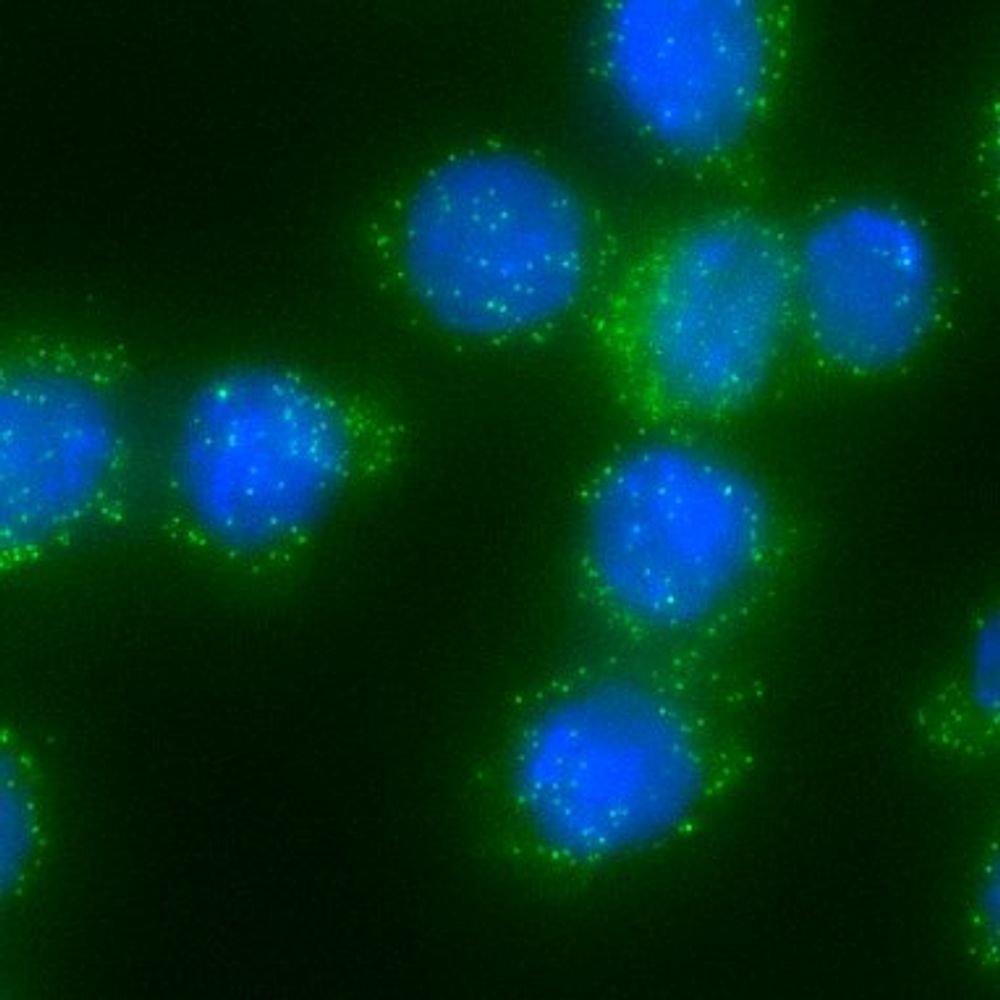
Overview of Integrated Robotic Imaging and Sequencing, IRIS
We just released IRIS (7+yrs project), a tech we believe will transform cell biology by pairing high-resolution cell images with matched #scRNAseq, letting us interpret cellular form by its molecular ground truth.
Huge tx to @JohannesBues, @JoernPezoldt, @CamilleLambert et al.
shorturl.at/zgY8Z
02.12.2025 14:17 — 👍 47 🔁 14 💬 2 📌 2
Congrats Elphège and Karissa. Very beautiful and important additional data in the final version!
28.11.2025 11:56 — 👍 1 🔁 0 💬 0 📌 0
And this is a fantastic result too
28.11.2025 11:54 — 👍 6 🔁 0 💬 0 📌 0

(23) If you move SRR2 further than ~20kb from the Sox2 promoter, it cannot support cohesin-independent action of the distal enhancer.
~20kb was the cuttof we say at Car2 for cohesin-independent action of enhancers. Makes sense!
27.11.2025 21:58 — 👍 2 🔁 1 💬 1 📌 1
This is a fantastic result!
28.11.2025 11:54 — 👍 1 🔁 0 💬 0 📌 0

The SMC Hinge is a Selective Gate for Obstacle Bypass - Nature Communications
SMC complexes are ring-shaped motors that fold DNA by extruding loops, but how they navigate large DNA obstacles is unclear. Here, Liu et al., show that SMC complexes bypass obstacles by threading obs...
The hinge bypass gate paper is finally out! doi.org/10.1038/s414...
We show how loop-extruding SMC complexes can maintain DNA entrapment while bypassing obstacles on DNA — including transcription machinery & potentially other SMCs.
A lucky convergence of 3 projects lead to the initial discovery!
25.11.2025 15:26 — 👍 30 🔁 15 💬 1 📌 2

Poster advertising the 6th X-inactivation meeting from Oct 19-23 2026 in Sapporo, Japan. The organizers are Asato Kuriowa, Edda Schulz, Ikuhiro Okamoto, Rafael Galupa, Takashi Sado, Mitinori Saitou.
📣 SAVE THE DATE
Next X-inactivation meeting in Sapporo, Japan, 19-23 October 2026. Visit x-inactivation-meeting.org to join our mailing list. 🧬 speakers @dandergassen.bsky.social @marnieblewitt.bsky.social @heard65.bsky.social @crougeulle.bsky.social @sexchrlab.bsky.social @zhouqi1982.bsky.social
17.11.2025 11:38 — 👍 36 🔁 18 💬 0 📌 0
Thanks for pointing this out - #3 is correct but in #7 there was a mistake: correct version is
"7/Loop extrusion-driven, long-lived encounters are longer than random collisions (up to tens of seconds vs. few seconds at most) but are much shorter than CTCF-anchored loops (which last 10-30 MIN)."
24.10.2025 17:34 — 👍 1 🔁 0 💬 1 📌 0
Our collab w. V Goel, @nicholas-aboreden.bsky.social , J Jusuf, G Blobel, L Mirny, @irate-physicist.bsky.social out in @natsmb.nature.com www.nature.com/articles/s41...
Was co-submitted with @allanaschooley.bsky.social @jobdekker.bsky.social whose paper should also come out soon
Brief thread 👇
20.10.2025 18:36 — 👍 55 🔁 18 💬 2 📌 2

Characterization of induced cohesin loop extrusion trajectories in living cells
Nature Genetics - This study introduces a system called TArgeted Cohesin Loader (TACL) that recruits cohesin complexes at defined genomic regions and induces loop extrusion events in living cells,...
The TArgeted Cohesin Loader (TACL) paper was just published. Happy that we were able to contribute to this really exciting project!
If you want to learn how targeting cohesin to defined loci in the genome affects the local chromatin environment and transcription, look no further!
rdcu.be/eLiT5
16.10.2025 20:17 — 👍 54 🔁 20 💬 0 📌 0

We are recuiting two new Associate Professors here in Oxford Biochemistry. Come join us! Reach out to me if you have any questions. Please repost! tinyurl.com/mr3m7bd3
17.10.2025 14:56 — 👍 77 🔁 98 💬 0 📌 1
Join us at the Hubrecht Institute in Utrecht to start your research group! Fantastic scientific environment, strong support, and nice colleagues :) Apply and please share broadly in your network.
28.09.2025 09:43 — 👍 16 🔁 23 💬 2 📌 0
The latest work from ours and @vram142.bsky.social lab is out! True teamwork to visualize nascent chromatin with strand resolution, using a fully reconstituted system. Very proud of superstar-PhD student Bruna, and @palindromephd.bsky.social. Learning so much from Vijay’s amazing technologies! RT
21.09.2025 07:12 — 👍 37 🔁 12 💬 2 📌 0
Unfortunately we don't have any data or models to answer your question. If nuclear shape changes affect levels of cofactors that determine extrusion rates (NIPBL,PDS5A/B) this could in principle affect extrusion velocity, but I can't personally point you to any data that support this scenario
26.09.2025 18:27 — 👍 0 🔁 0 💬 1 📌 0
The 'encounter radius' is an arbitrary physical range, which we vary systematically - and show that its particular choice doesn't affect any of the conclusions.
26.09.2025 17:49 — 👍 1 🔁 0 💬 1 📌 0
As you will see in the manuscript, the only condition for two loci to be 'caught' in an extrusion-driven encounter is that cohesin starts extruding approx. midway between their genomic positions (plus of course that their genomic distance is not much larger than cohesin's processivity)
26.09.2025 17:47 — 👍 1 🔁 0 💬 1 📌 0
Thanks for the kind words Anders - indeed this is a very much overlooked issue that also affects interpretation of DNA FISH data. Even small amounts of error make determination of 'interactions' unreliable, as your lab also pointed out earlier e.g. in www.sciencedirect.com/science/arti...
26.09.2025 17:43 — 👍 4 🔁 0 💬 0 📌 0
We do not impose any specific range, but rather compute (or measure) any events where the two loci spend time within an arbitrary encounter range (R_e in the manuscript), either mediated by random fluctuations or by loop extrusion...
26.09.2025 07:16 — 👍 2 🔁 0 💬 1 📌 0
you guys are too fast :)
24.09.2025 21:45 — 👍 2 🔁 0 💬 0 📌 0
18/ At large genomic distance, long encounters (=the important ones) are only due to loop extrusion; but at short genomic distance they can also be due to random collisions. This is why enhancers are no sensitive to loss of extrusion when they are close, but sensitive when they are far!
24.09.2025 21:45 — 👍 2 🔁 1 💬 1 📌 0

17/ ... and nicely predicts @elphegenoralab.bsky.social @karissalhansen.bsky.social's data at the Car2 locus www.biorxiv.org/content/10.1..., using changes in cohesin occupancy and extrusion velocity measured with @gfudenberg.bsky.social in www.biorxiv.org/content/10.1...:
24.09.2025 21:45 — 👍 3 🔁 0 💬 1 📌 0
16/ Finally, this simple model also predicts and explains why, at least within simple genomic locations devoid of further confounding effects, depletion of cohesin or its loading and elongation factor NIPBL affects enhancer function at large, but nor short genomic distance...
24.09.2025 21:45 — 👍 2 🔁 0 💬 1 📌 0

15/ This happens because by selecting longer and longer encounters, one selects more and more loop-extrusion-driven events, the probability of which is exponential as a function of genomic distance!
24.09.2025 21:45 — 👍 2 🔁 0 💬 1 📌 0

14/ This simple model also predicts that transcription levels should decrease exponentially as a function of genomic distance between an enhancer and promoter, exactly as we verified using data from our previous work!
24.09.2025 21:45 — 👍 2 🔁 0 💬 1 📌 0
Research group at SISSA / Trieste. Interested in stochastic processes in biology & biophysics. Posts by C. Zechner.
Research Group Leader @MRC_LMS @imperialcollege
How are biological forms encoded? Group leader at IBDM Marseille, director of Turing Center for living systems (CENTURI). Professor at Collège de France, Paris: Dynamics of living systems
DevBiologist ❤️ gene regulation. Postdoc on mouse limbs in Mundlos Lab MPI for Molecular Genetics. Previously PhD on ascidian enhancer-evo in Lemaire Lab CNRS-CRBM | she/her
Group leader at Institut Jacques Monod. When I say "epigenetics" what I really mean is "DNA methylation".
www.maximgreenberglab.com
🇧🇷 Postdoc @ MPI Molecular Genetics (Berlin) interested in genetics, synthetic biology and embryo development. Previously @ EMBL and UFRJ.
PhD student at the Netherlands Cancer Institute || gene regulation - epigenetics - genome organization || she/her
We are a group at the Center for Molecular Biology (ZMBH) at Heidelberg University that strive to understand the regulatory principals that govern gene expression in health and disease
Asst Prof at ISTAustria. Lover of tiny things including proteins, tardigrades, seashells & round furry animals.
Genomics, Machine Learning, Statistics, Big Data and Football (Soccer, GGMU)
An international online community and seminar series about chromatin and gene regulation.
Join the community on Discord: http://discord.gg/dXqT89r
Subscribe to our mailing list: https://stats.sender.net/forms/bmVp3d/view
Chromatin and transcription aficionado. Postdoc in Bickmore lab.
Assistant Professor of Cell Biology at Harvard Medical School | HHMI Freeman Hrabowski Scholar | Nexus of chromatin, transcription, replication, and epigenetics. farnunglab.com
Assistant Professor at UTSW studying nuclear organization by nuclear condensates
Researcher of chromatin, transcription, DNA repair, splicing & actin. Mostly microscopy. @unibirmingham
The Nogales Lab @ UC Berkeley. #CryoEM #Transcription #Microtubules #ChromatinModification Bluesky account managed by lab members.
Postdoc with @Giacomo_Cavalli, former PhD student in the @marstonlab.bsky.social. HFSP Fellow, EMBO Fellow. 3D genome enthusiast!
Deputy Director at WEHI, Melbourne Australia.
Lab head studying epigenetic control, in the context of X inactivation, genomic imprinting, SMCHD1, Prader Willi Syndrome, FSHD.
Mum, wife, beach lover.