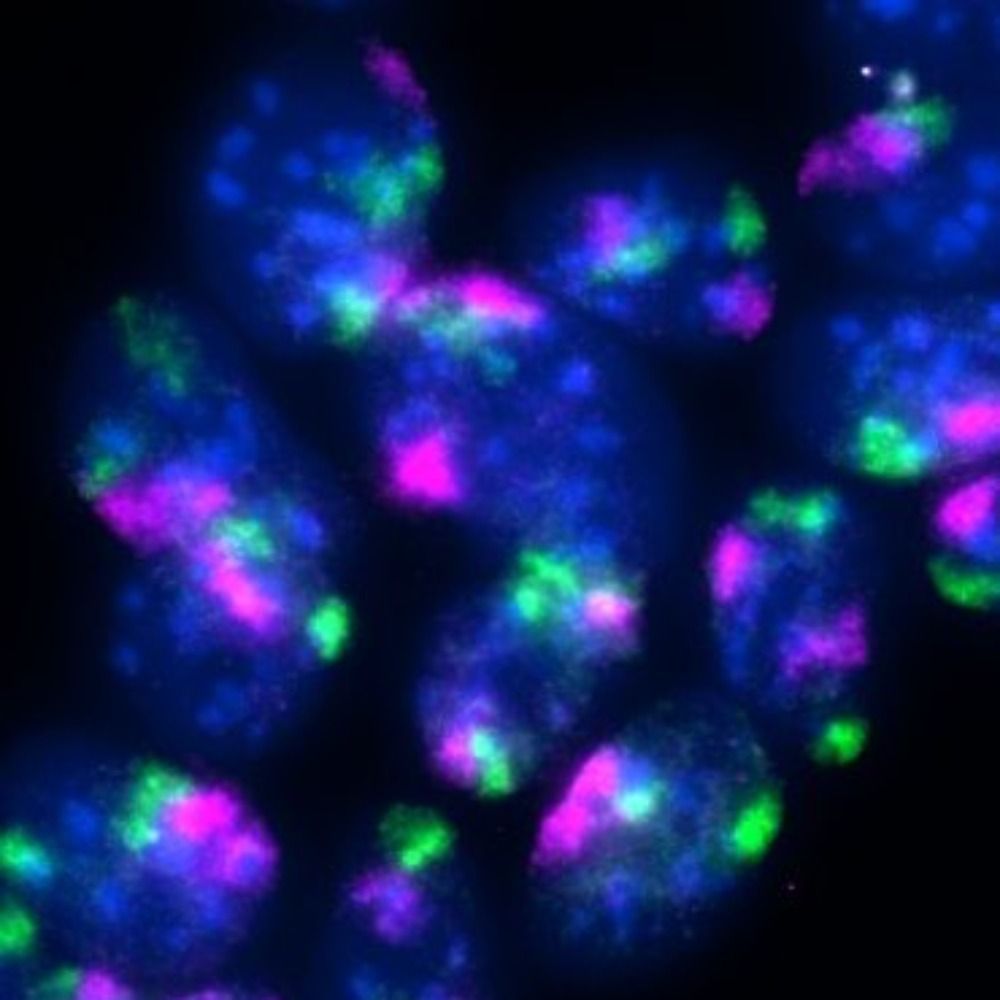
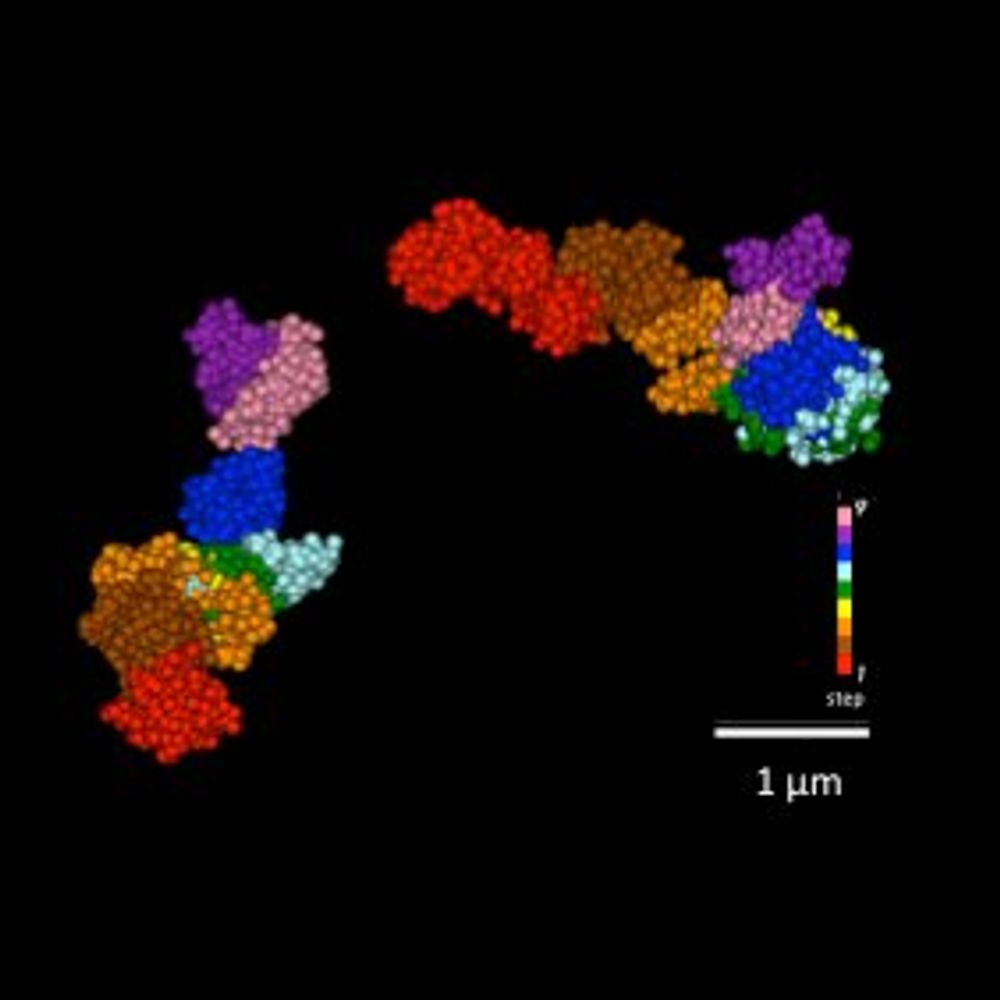
10/12
Our model:
🔹 Loop-forming cohesin sets a megabase-scale search radius within the TAD
🔹 Cohesive cohesin forms a local clamp that tethers DNA ends to the sister chromatid
Together, they focus RAD51 sampling and enable accurate repair!
04.12.2025 20:17 — 👍 2 🔁 2 💬 1 📌 0
YouTube video by Sciences de la vie - Collège de France
L'inactivation du chromosome X (12) - Edith Heard (2024-2025)
(29) PS: here is a link to a public presentation of this work on enhancer biology
m.youtube.com/watch?v=8OdG...
Courtesy @college-de-france.fr
04.12.2025 13:55 — 👍 1 🔁 0 💬 0 📌 0

📣 I hereby make my Bluesky debut to announce that our work linking DNA binding affinities and kinetics 𝘪𝘯 𝘷𝘪𝘵𝘳𝘰 and 𝘪𝘯 𝘷𝘪𝘷𝘰 for the human transcription factor KLF1 just got published in Cell! @cp-cell.bsky.social
www.cell.com/cell/fulltex...
Key findings in a thread (1/6):
27.11.2025 13:17 — 👍 98 🔁 33 💬 3 📌 2
🧩 What breaks when you remove DNA loop extrusion?
Gastruloids show it: not lineage decisions… but morphogenesis.
That’s our contribution to www.science.org/doi/10.1126/...
Huge thanks to the amazing @karissalhansen.bsky.social + @elphegenoralab.bsky.social for leading this fantastic work 🙌
28.11.2025 08:57 — 👍 7 🔁 2 💬 0 📌 0

(27) We could recreate CTCF insulator bypass at the Car2 locus by introducing SRR2
🤯
> How many TAD boundaries throughout the genome get bypassed by enhancers through this synergy mechanism? 👀 👀 👀
27.11.2025 21:58 — 👍 1 🔁 0 💬 1 📌 0

(25) Putting SRR2 Car2 makes the locus less reliant on cohesin loop extrusion
> so proximal and distal enhancers seem to synergize in a pretty degenerate way (read: it's not locus-specific, you can mix and match regulatory elements, to some extent)
27.11.2025 21:58 — 👍 2 🔁 0 💬 1 📌 0

(24) Using 4C we did not find evidence regulatory element synergy works by maintaining long-range chromosome folding independently of cohesin 😦
It also does not operate LDB1 here ❌
> molecular mechanism of enhancer synergy still to be investigated (get in touch!) 🔍
27.11.2025 21:58 — 👍 1 🔁 0 💬 1 📌 0

(23) If you move SRR2 further than ~20kb from the Sox2 promoter, it cannot support cohesin-independent action of the distal enhancer.
~20kb was the cuttof we say at Car2 for cohesin-independent action of enhancers. Makes sense!
27.11.2025 21:58 — 👍 2 🔁 1 💬 1 📌 1

(22) Strikingly, you can replace SRR2 with other weak enhancers to restore cohesin-independence at Sox2
> SRR2 is not special
> proximal and distal elements likely generally synergize in a cohesin-independent way
(thank you @chribue.bsky.social for highlighting the Map4k3 E2 enhancer!)
27.11.2025 21:58 — 👍 3 🔁 0 💬 1 📌 1
(21) ✨New data in the revised version ✨:
a) is SRR2 special in how it bolsters the action of the distal Soz2 enhancer independently of cohesin?
b) How does it work?
c) can we transplant it to change the cohesin-dependence a locus (e.g. Car2?)
27.11.2025 21:58 — 👍 1 🔁 0 💬 1 📌 0
(20) Additional authors on bluesky
@rinishah.bsky.social
@bffswithbiology.bsky.social
27.11.2025 21:58 — 👍 0 🔁 0 💬 1 📌 0
(19) This was a heroic effort by graduate student
@karissalhansen.bsky.social with the help of @annieadachi.bsky.social , and many wonderful collaborators in our lab and in
@dewitlab.bsky.social
@gfudenberg.bsky.social
@robertblelloch.bsky.social
groups
27.11.2025 21:58 — 👍 1 🔁 0 💬 1 📌 0
(18) How does the SRR2 work to boost the effect of distal enhancers? Condensate biophysics? Unknown looping factors? 🤷
How can we hunt for similar elements across the genome & across cell types? 🧪🧬❓
We would love to know – check out the paper for some discussion.
27.11.2025 21:58 — 👍 0 🔁 0 💬 1 📌 0

(17) So it’s not that complicated in the end:
If you rely on an enhancer, you will need cohesin extrusion beyond ~18kb.
UNLESS you have a promoter-proximal regulatory element that can synergize with your distal enhancer – which happens independently of
27.11.2025 21:58 — 👍 0 🔁 0 💬 1 📌 0

(16) Why? Quite simply, you have now removed the two redundant axes that support long-range enhancer action at Sox2.
27.11.2025 21:58 — 👍 0 🔁 0 💬 1 📌 0

(15) That’s because SRR2 is there to support cohesin-independent regulatory communication:
Delete SRR2 when extrusion is blocked by the CTCF sites, and now they insulate very well.
27.11.2025 21:58 — 👍 0 🔁 0 💬 1 📌 0

(14) Others already noticed that distal enhancers can still activates Sox2 even if you put really strong CTCF sites in between.
We see that’s true even for sites that insulate *very well* at the Car2 locus.
Why is CTCF insulation not working well at Sox2?
27.11.2025 21:58 — 👍 0 🔁 0 💬 1 📌 0
13) Are there other contexts where the cohesin-independent synergy between SRR2 and distal enhancers is at play?
Yes, when we block extrusion with strong CTCF sites.
🛑🚧🛑
27.11.2025 21:58 — 👍 0 🔁 0 💬 1 📌 0

(12) So SRR2 mediates a cohesin-independent mechanism to support the communication of Sox2 with its distal enhancer 100kb away.
For Sox2, this regulatory axis and loop extrusion are redundant: that’s why you need to disable both to see a transcriptional effect.
27.11.2025 21:58 — 👍 0 🔁 0 💬 1 📌 0

(10) Wait. How can Sox2 work without extrusion? Its enhancer is 100kb away – that’s very far.
Loooots of detective work later 🕵️♀️ the answer is crystal clear: it’s all about an inconspicuous genetic element 3kb downstream of the promotor called SRR2.😲
27.11.2025 21:58 — 👍 0 🔁 0 💬 1 📌 0

(9) What is we drive Car2 with a crazy strong enhancer?
Let’s grab the Sox2 super-enhancer, since we know it does not really need cohesin to work at Sox2.
Same cut off: 18kb.
So your host locus, not your enhancer, decides if you need cohesin or not for distal regulation.
27.11.2025 21:58 — 👍 0 🔁 0 💬 1 📌 0

(8) By relocating enhancers closer and closer to the Car2 promoter we can render it completely resistant to inhibiting extrusion, although it is normally very dependent on extrusion
The cut off: 18kb. Above that the Car2 enhancers need extrusion to work.
27.11.2025 21:58 — 👍 0 🔁 0 💬 1 📌 0

(7) Do enhancers need cohesin to work then?
Some yes (e.g. at the Car2 locus)
Some not really (e.g. everyone’s favorite Sox2)
Why is that?
27.11.2025 21:58 — 👍 0 🔁 0 💬 1 📌 0

(6) Back to the gene loci that are dysregulated: why are some but not all genes messed up?
Smit Kadvani and
@gfudenberg.bsky.social
noticed that down-regulated genes lie in enhancer-rich chromosome neighborhoods
That makes sense with the tissue-specific dysregulations.
27.11.2025 21:58 — 👍 0 🔁 0 💬 1 📌 0
Aging lab @Stanford. Our interests include mechanisms of aging, brain aging and rejuvenation, neural stem cell aging, genetics of lifespan and suspended animation in killifish
Independent Group Leader at the MPI-IE passionate about mechanisms governing embryonic development with a focus on transposable elements🧬 Postdoc @MundlosLab 🇩🇪| PhD from @Bourchis lab 🇫🇷| She/her
https://www.julianeglaserlab.com/
Studying specializations and vulnerabilities of human brain development
DNA Topology Lab @ Department of Biochemistry
University of Oxford
"We adore chaos because we love to produce order." -M. C. Escher
EPIGENETIC HULK SMASH PUNY GENOME. MAKE GENOME GO. LOCATION: NOT CENTROMERE, THAT FOR SURE
Scientist at IMP in Vienna. Excited about gene expression regulation and its encoding in our genomes - enhancers, transcription factors, co-factors, silencers, AI.
Biologist especially interested in cilia, signaling and development!
#Chromatin biologist/biophysicist @NIG & SOKENDAI in Japan.
Chromatin is very dynamic and flexible, but NOT regular!!! 🧪🧬🔬
My career and work: http://bit.ly/2CuF4L5
Lab HP: https://bit.ly/3F1a8nk
YouTube Seminar: https://bit.ly/4fYZiOj
NIH-F32 Postdoctoral Fellow @ChoryLab @DukeCAGT
Chromatin, SynBio, Automation
PhD @UCSF | #FirstGen | Nice to meet you friend☀️🌲😊
Just another LLM. Tweets do not necessarily reflect the views of people in my lab or even my own views last week. http://rajlab.seas.upenn.edu https://rajlaboratory.blogspot.com
Postdoc in Boettiger lab @Stanford
https://pownall-lab.org | Sandler Fellow at UCSF | chromatin, transcription, microscopy (ChromExM), development 🐟🧬🔬
Assistant Professor at UTSW studying nuclear organization by nuclear condensates
We are the Ting Wu Lab @ HMS. Our laboratory studies how chromosome behavior and positioning influence genome function, with implications for gene regulation, genome stability, and disease.
Managed by WuLab members
https://www.transvection.org/home
EMBO postdoctoral fellow at LMS - MRC in Lymphocyte Development group | Interested in genome organisation and SMC complexes 🪡
Department of Genetics, Institute for Immunology and Immune Health, Epigenetics Institute, University of Pennsylvania
https://vahedilab.com/ https://www.instagram.com/vahedilaboratory/
Assistant Prof @PittBioSci studying transcription dynamics and chromatin biology | passionate about equity in STEM | she/her